医学及细胞生物学研究

近日,中国药科大学马广勇研究员团队在《Nature Communications》发表题为 "HTLV-1 subverts the innate immune effector gene IRF7 by viral HBZ protein for oncogenesis" 的研究论文,第一作者为袁小艺博士。该研究围绕 HTLV-1 相关成人 T 细胞白血病/淋巴瘤(ATLL)发生机制,系统揭示了病毒癌蛋白 HBZ 如何"劫持"先天免疫关键转录因子 IRF7,使其脱离传统抗病毒 IFN-I 通路,转而通过 IRF7–STAT5B 轴促进肿瘤细胞增殖与存活。
本研究中,单细胞转录组测序由烈冰生物采用自研的 NovelCyto 单细胞平台完成;单细胞数据分析由作者在烈冰生物自研的 CytoNavigator 单细胞数据分析系统协助中完成。
HTLV-1 是目前明确具有致癌性的经典人类逆转录病毒之一,可导致成人 T 细胞白血病/淋巴瘤(ATLL)。ATLL 具有侵袭性强、治疗选择有限、预后较差等特点,其发生发展与病毒编码蛋白 Tax 和 HBZ 密切相关。其中,Tax 具有较强的转录激活能力,但在体内感染细胞尤其是 ATLL 患者中往往表达不稳定;相比之下,HBZ 在 HTLV-1 感染个体中持续表达,被认为是维持感染细胞生存和促进肿瘤发生的重要病毒因子。
IRF7 是先天免疫反应中的核心转录因子,通常与 IRF3 共同诱导 I 型干扰素(IFN-I)产生,进而启动 JAK/STAT 介导的抗病毒反应。传统观点认为,病毒往往通过抑制 IRF7 来逃避免疫识别;而该研究提出了一个更具颠覆性的机制:HTLV-1 并非简单关闭 IRF7,而是通过 HBZ 上调并激活 IRF7,同时干扰 IRF7–IRF3 介导的经典 IFN-I 抗病毒功能,使 IRF7 转变为支持 ATLL 细胞生长的"致癌因子"。
|
模块 |
研究内容 |
|
体内模型 |
斑马鱼胚胎模型;通过注射 HTLV-1 感染性克隆、HBZ/Tax 表达载体或 mRNA,观察胚胎发育异常及存活变化。 |
|
转录组筛选 |
bulk RNA-seq 评估 HBZ 诱导的全转录改变,提示先天免疫相关 PRR 通路被显著激活。 |
|
单细胞解析 |
对 3 dpf 斑马鱼胚胎头部组织进行 scRNA-seq,单细胞测序由烈冰生物完成;作者使用 CytoNavigator 单细胞数据分析系统进行细胞聚类、细胞类型鉴定及基因集富集分析等。 |
|
临床数据验证 |
结合 ATLL 患者公开 scRNA-seq 数据集(PRJEB47382)验证关键通路及 IRF7 表达变化。 |
|
机制与功能验证 |
CUT&Tag、双荧光素酶、Co-IP、GST pull-down、CRISPR/shRNA 干预、细胞增殖/凋亡实验及小鼠移植瘤模型等。 |
━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━
研究首先利用斑马鱼胚胎模型快速评估 HTLV-1 病毒基因的体内效应。斑马鱼与人类基因高度保守,且兼具先天免疫与适应性免疫系统,适合用于快速评估病毒蛋白对发育、免疫及血液系统相关表型的影响。结果显示,注射 HTLV-1 感染性克隆可导致明显的胚胎发育异常,包括脊柱弯曲和心包水肿,并增加胚胎死亡率。进一步基因功能拆解发现,敲除 HBZ 可显著缓解上述表型,而敲除 Tax 并不能产生类似保护效应,提示 HBZ 是 HTLV-1 诱导斑马鱼发育异常的主要驱动因子。
在 HBZ 的不同功能形式中,研究进一步证明其致病效应仅依赖 HBZ 蛋白形式,RNA 形式在该实验体系下未表现出相同致病作用;其中 activation domain(AD)和 bZIP 结构域对该过程至关重要。结合 bulk RNA-seq 分析,HBZ 表达可引起大量基因表达重塑,并显著激活 TLR、CLR、RLR、NLR 等先天免疫识别通路。
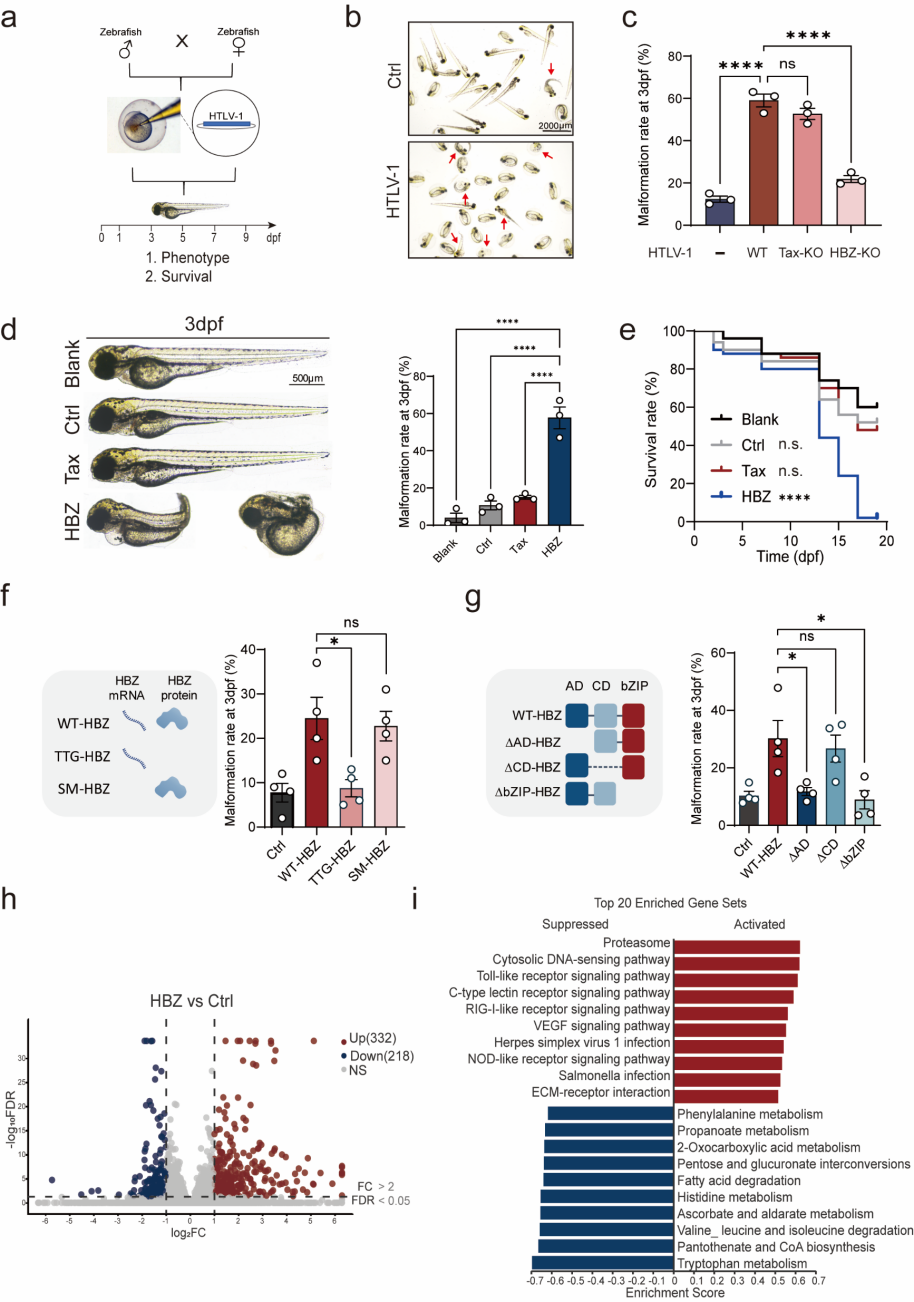
[图1] HBZ 而非 Tax 是 HTLV-1 诱导斑马鱼胚胎发育异常的关键病毒因子,并伴随先天免疫相关通路激活
为进一步从细胞异质性层面解析 HBZ 对胚胎组织的影响,研究者对 3 dpf 斑马鱼胚胎样本开展单细胞转录组测序,最终鉴定出 27 个细胞群。单细胞结果显示,HBZ 表达主要分布于免疫相关细胞,包括造血/淋巴样细胞,同时也可见于部分上皮细胞。
与 bulk RNA-seq 结果一致,单细胞层面可观察到 CLR、NLR、TLR 等先天免疫识别通路被激活。更重要的是,研究者进一步结合 ATLL 患者公开 scRNA-seq 数据集,发现 NLR/RLR 等先天免疫相关通路在临床 ATLL 样本中同样处于活化状态。跨模型对照分析最终锁定 IRF7:该基因同时在 HBZ 表达的斑马鱼胚胎和 ATLL 临床样本中上调。
这一部分正体现了单细胞测序的核心优势:scRNA-seq 并非仅用于"画图分群",而是帮助研究者从复杂组织状态中定位 HBZ 主要作用的细胞区域和共同上调的关键免疫转录因子,为后续机制验证提供了清晰方向。
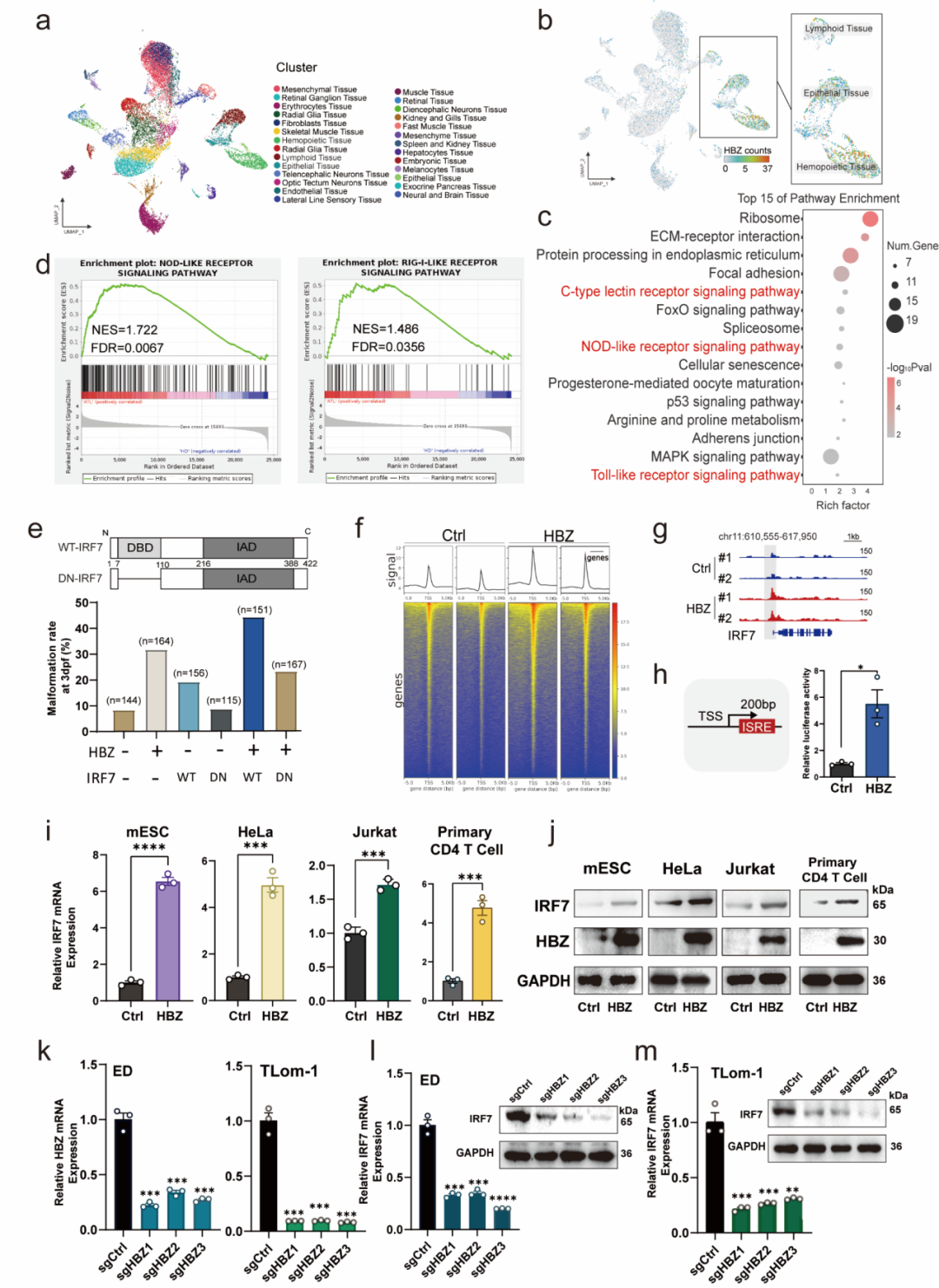
[图2] 单细胞转录组解析显示 HBZ 表达相关细胞群及先天免疫识别通路活化,并进一步锁定 IRF7 为共同上调的关键因子
研究者证实 HBZ 直接调控 IRF7:斑马鱼胚胎注射 IRF7 mRNA,可重现 HBZ 诱导的发育异常表型;相反,表达 dominant-negative IRF7 可部分挽救 HBZ 诱导的病理表型,提示 IRF7 是 HBZ 下游重要功能节点。
机制上,CUT&Tag 结果显示 HBZ 可富集于宿主基因启动区附近,并在 IRF7 启动子上游区域出现明显富集;双荧光素酶实验进一步证实 HBZ 能增强 IRF7 启动子活性。与此同时,HBZ 可在小鼠 ESC、Jurkat、HeLa 以及原代人 CD4 T 细胞中上调 IRF7 的 mRNA 和蛋白表达;在 ATLL 细胞系中敲低内源性 HBZ,则会降低 IRF7 表达。
除转录上调外,HBZ 还可促进 IRF7 Ser477 位点磷酸化并驱动其核转位。通过激酶抑制剂和蛋白检测,研究进一步指出 TBK1 是 HBZ 介导 IRF7 磷酸化的重要上游激酶。由此,HBZ 通过"提高 IRF7 表达量 + 促进 IRF7 激活状态"两条路径,增强 IRF7 功能。
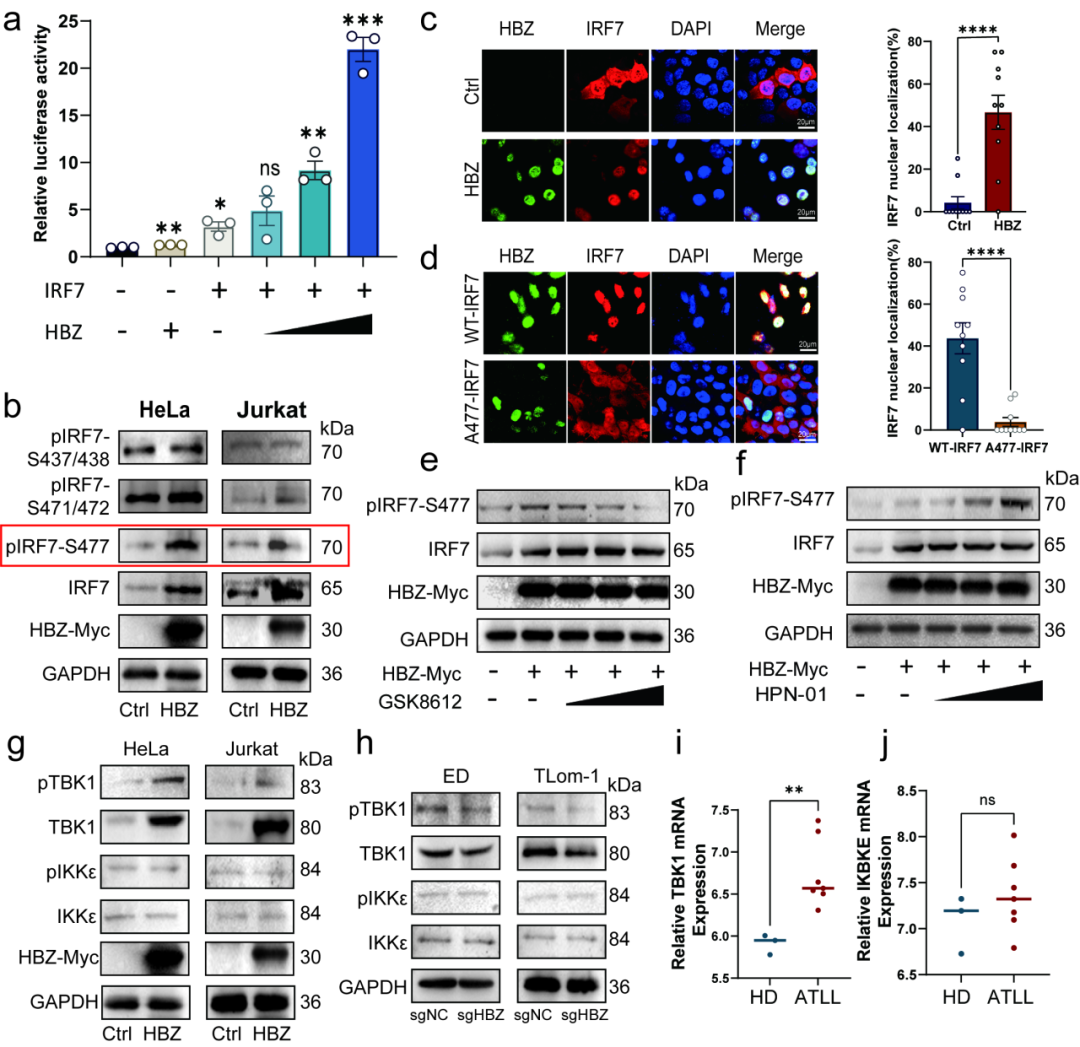
[图3] HBZ 通过促进 IRF7 磷酸化、核转位及 TBK1 激活,增强 IRF7 的功能状态
按照经典免疫学逻辑,IRF7 被激活后应诱导 IFN-I 表达并启动抗病毒反应。然而该研究发现,在 HBZ 表达的斑马鱼胚胎和 ATLL 临床样本中,尽管 IRF7 显著上调,IFN-I 通路关键基因并未同步激活。
进一步实验表明,poly(I:C) 可在细胞中诱导 IFN-α 产生,但 HBZ 的引入会削弱这一反应。结构预测、Co-IP 与 GST pull-down 结果共同显示,HBZ 可与 IRF7 直接相互作用,其中 HBZ 的 bZIP 结构域对该相互作用十分关键。更重要的是,HBZ 能干扰 IRF7 与 IRF3 的相互作用,阻碍二者形成有效的抗病毒转录复合体,从而抑制其共同诱导 ISRE 活性的能力。
也就是说,HTLV-1 并不是简单"关闭"IRF7,而是更精细地重塑了 IRF7 的功能方向:一方面保留并增强 IRF7 的表达和核内活性;另一方面干扰 IRF7 与 IRF3 的相互作用,阻碍抗病毒转录复合体形成,使 IRF7 无法有效启动 IFN-I 抗病毒程序。
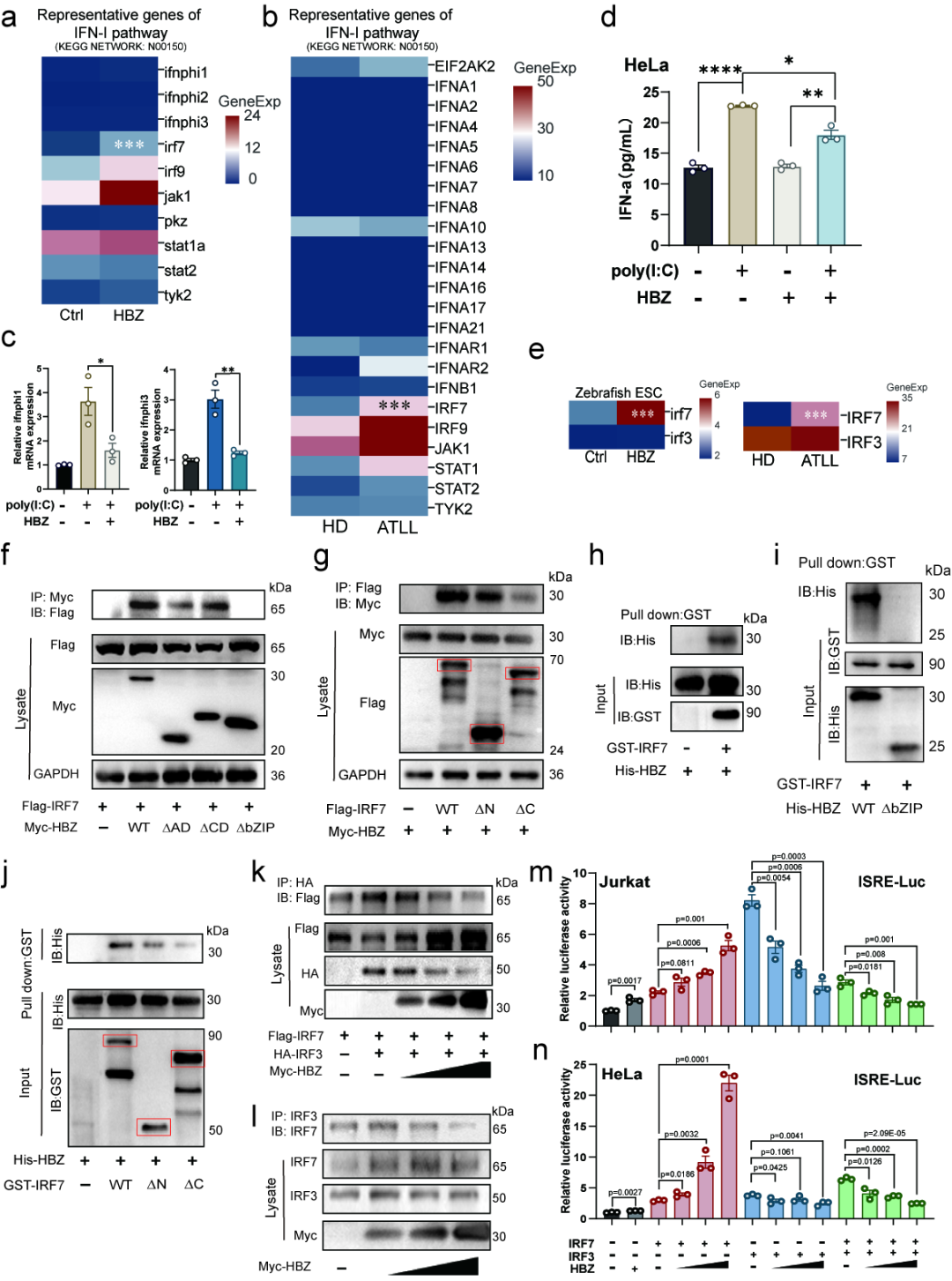
[图4] HBZ 与 IRF7 直接互作并干扰 IRF7–IRF3 相互作用,使 IRF7 脱离经典 IFN-I 抗病毒通路
在 ATLL 细胞系 ED 和 TL-Om1 中,shRNA 介导的 IRF7 敲低显著抑制细胞增殖,并增加细胞凋亡。研究者进一步建立免疫缺陷小鼠移植瘤模型,发现 IRF7 敲低可明显延缓肿瘤生长、降低肿瘤体积;而 IRF7 回补实验能够恢复细胞增殖和肿瘤生长能力,从功能上验证 IRF7 在 ATLL 中具有促肿瘤作用。
研究还通过分子对接筛选到潜在 IRF7 小分子抑制剂 Ditercalinium。Ditercalinium 处理可降低 IRF7 蛋白水平,并抑制 ATLL 细胞系增殖。这一结果提示,IRF7 不仅是机制链条中的关键节点,也可能为 ATLL 后续靶向干预研究提供新的候选方向。
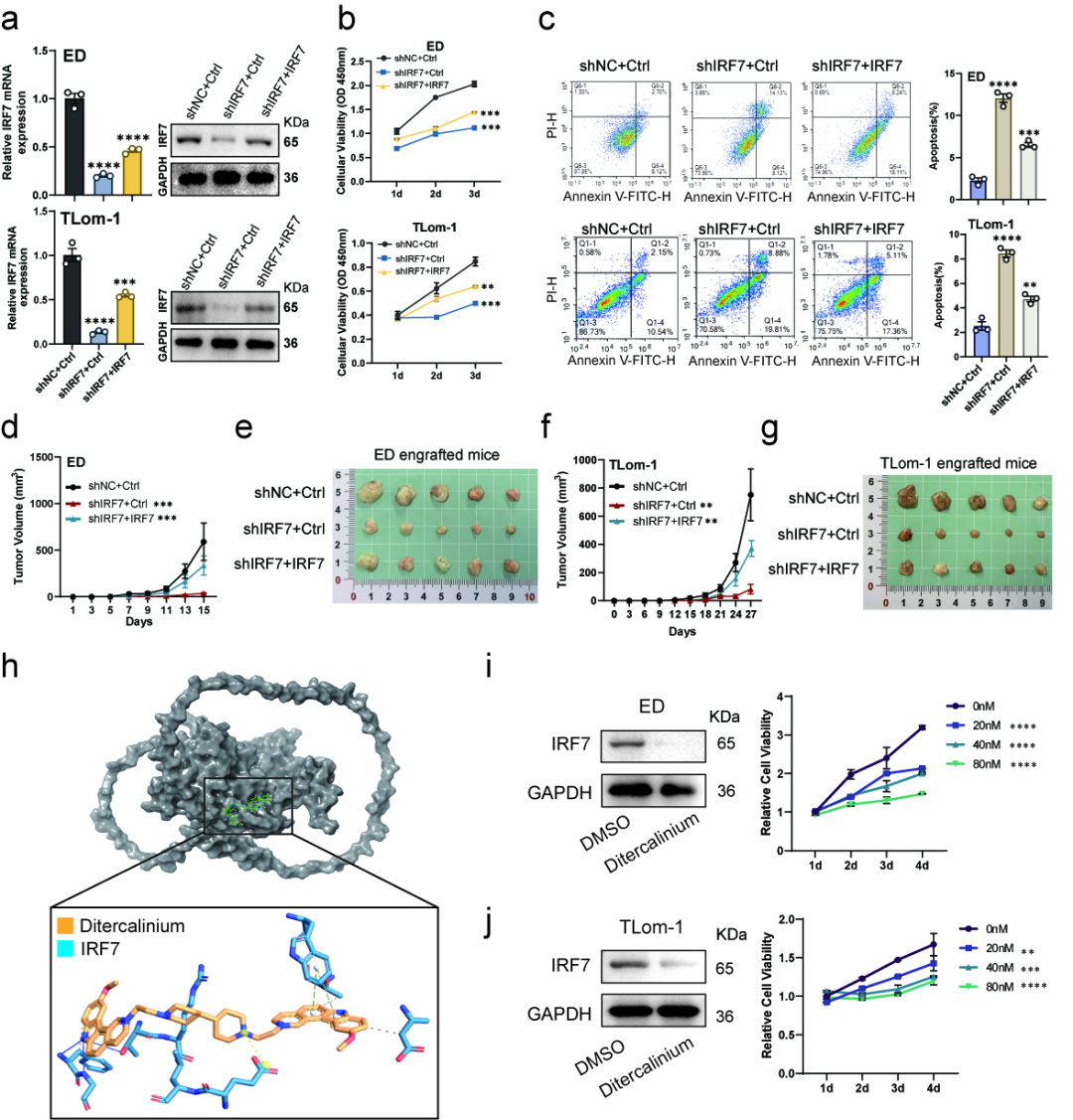
[图5] IRF7 在 ATLL 中表现出促肿瘤功能,敲低 IRF7 可抑制细胞增殖、增加凋亡并降低移植瘤生长
为解释 IRF7 如何促进 ATLL 细胞生长,研究者在 IRF7 敲低后的 ATLL 细胞系中进行转录组分析,并筛选出在 IRF7 敲低后下调、同时在 ATLL 患者样本中上调的共同靶基因。其中,STAT5B 作为 JAK/STAT 通路中的关键转录因子被重点关注。需要注意的是,STAT5B 功能获得性突变在 T-ALL、儿童 T-ALL、NK-LGLL 等造血恶性肿瘤中反复出现,但该研究指出 ATLL 患者中并未发现这类突变;本研究关注的是 STAT5B 在 ATLL 中的高表达及功能激活。
后续实验显示,IRF7 敲低可降低 STAT5B 的 mRNA 和蛋白水平;CUT&Tag 结果显示 IRF7 在 STAT5B 基因座多个区域存在富集,提示其可能直接调控 STAT5B 转录。此外,IRF7 还可与 STAT5B 发生蛋白相互作用,并增强 STAT5B 的转录激活能力。功能上,STAT5B 回补能够挽救 IRF7 敲低导致的细胞增殖抑制,说明 STAT5B 是 IRF7 促 ATLL 细胞生长的重要下游效应分子。
最终,研究提出了 HBZ–IRF7–STAT5B 致癌轴:HTLV-1 编码的 HBZ 上调并激活 IRF7;HBZ 同时干扰 IRF7–IRF3 介导的经典 IFN-I 抗病毒功能;被"重新定向"的 IRF7 进一步促进 STAT5B 表达与活性,推动 ATLL 细胞存活和增殖。
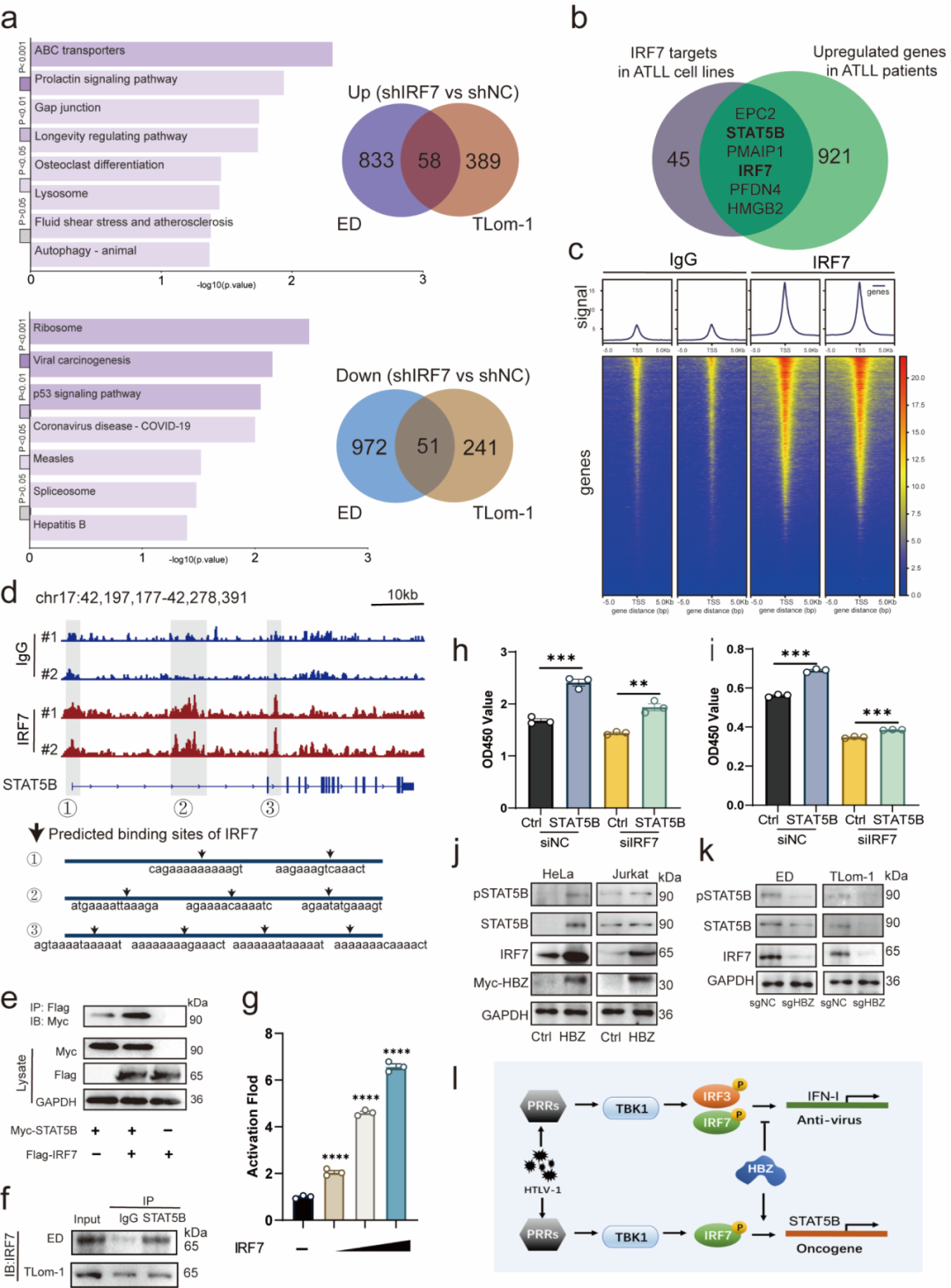
[图6] IRF7 通过调控并激活 STAT5B 形成 HBZ–IRF7–STAT5B 致癌轴,推动 ATLL 细胞存活与增殖
━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━
该研究重新定义了 IRF7 在 HTLV-1 相关 T 细胞恶性肿瘤中的功能定位,提出 IRF7 可被病毒癌蛋白 HBZ 重新编程为促肿瘤因子。
单细胞转录组帮助研究者在斑马鱼胚胎复杂细胞组成中解析 HBZ 作用细胞群及先天免疫识别通路活性,并通过与 ATLL 临床单细胞数据交叉验证,锁定 IRF7 这一核心共性分子。
研究从体内斑马鱼模型、细胞系、原代 CD4 T 细胞、ATLL 患者数据,到小鼠移植瘤模型,构建了从表型观察、单细胞发现、分子机制到功能验证的完整研究闭环。
IRF7 与 STAT5B 均表现出促 ATLL 细胞生长功能,相关抑制剂能够抑制 ATLL 细胞增殖,为 HTLV-1 相关 T 细胞恶性肿瘤的后续靶向干预研究提供了新的候选方向。
━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━
本研究揭示了 HTLV-1 促进 ATLL 发生的一种非常巧妙的病毒致癌策略:病毒蛋白 HBZ 并非单纯抑制先天免疫,而是选择性"劫持"IRF7。HBZ 一方面增强 IRF7 表达和磷酸化激活,另一方面干扰 IRF7 与 IRF3 的相互作用,阻碍经典抗病毒转录复合体形成,使其无法有效诱导 IFN-I;同时,IRF7 被重新导向 STAT5B 轴,进而促进 ATLL 细胞增殖、生存和肿瘤形成。
从技术路径看,单细胞转录组在该研究中承担了关键的机制发现角色:它帮助研究者从 HBZ 诱导的复杂组织反应中识别先天免疫通路活化和 IRF7 上调这一核心线索,并通过临床 ATLL 单细胞数据进行跨物种、跨模型验证。
━━━━━━━━━━━━━━━━━━━━━━━━━━━━━━
本研究中,斑马鱼胚胎单细胞转录组测序由烈冰生物采用自研的 NovelCyto 单细胞平台完成;单细胞数据分析由作者在烈冰生物自研的 CytoNavigator 系统协助中完成。
采用静态蜂巢板/微孔捕获体系,依靠重力沉降实现细胞捕获,反应过程柔和,对各类细胞无选择性偏好,尤其对粒系细胞、复杂样本和低浓度细胞悬液具有较好的适应性,能够为疾病机制研究产出稳定、高质量的单细胞转录组数据。
将单细胞数据分析流程拆解为清晰的模块化任务,0 代码,结合一键式分析、交互式可视化、结果简报生成和流程追溯功能,帮助研究者快速识别关键细胞亚群、差异基因及功能通路等信息,为研究者提供了系统化的数据分析支撑。

未来,烈冰生物也将持续以高质量单细胞测序与数据分析能力,助力更多疾病机制研究。
 联络我们
联络我们

021-52235399

tech@novelbrain.com

江西生产总部:江西省上饶市国际医疗旅游先行区上饶大道18号6号楼
上海运营总部:上海市闵行区江月路999号奇亚特中心22栋
北京分公司:北京市朝阳区红军营南路15号瑞普大厦C座
西安IT研发中心:西安市未央区赛高商务港2504
广州办事处:广州市越秀区中山三路33号中华国际中心B座3106
香港办事处:香港特别行政区沙田区科学园科技大道西16号
南京分公司:秦淮区正洪街18号东宇大厦C座17B
武汉办事处:湖北省武昌区中南路街民主路786号华银大厦21层4室
成都办事处:成都市成华区梦魔方广场A座2218
杭州办事处:杭州市拱墅区文晖路183号20楼007室











关注公众号


 生信分析平台
生信分析平台 实验进度查询
实验进度查询


